
Cấu trúc của một đoạn xoắn kép DNA.
Bạn đang đọc: DNA – Wikipedia tiếng Việt
DNA là phân tử mang thông tin di truyền quy định mọi hoạt động sống (sinh trưởng, phát triển và sinh sản) của các sinh vật và nhiều loài virus. Đây là từ viết tắt thuật ngữ tiếng Anh deoxyribonucleic acid, theo tiếng Việt gọi là acid deoxyribonucleic[1][2] (nguồn gốc từ tiếng Pháp: acide désoxyribonucléique, viết tắt: ADN).
DNA và RNA là những acid nucleic, cùng với protein, lipid và carbohydrat cao phân tử ( polysaccharide ) đều là những đại phân tử sinh học chính có vai trò quan trọng thiết yếu so với mọi dạng sống được biết đến. Phần lớn những phân tử DNA được cấu trúc từ hai mạch polymer sinh học xoắn đều quanh một trục tưởng tượng tạo thành chuỗi xoắn kép .
Hai mạch DNA này được gọi là các polynucleotide vì thành phần của chúng bao gồm các đơn phân nucleotide.[3][4] Mỗi nucleotide được cấu tạo từ một trong bốn loại nucleobase chứa nitơ—hoặc là cytosine (C), guanine (G), adenine (A), hay thymine (T)—liên kết với đường deoxyribose và một nhóm phosphat. Các nucleotide liên kết với nhau thành một mạch DNA bằng liên kết cộng hóa trị giữa phân tử đường của nucleotide với nhóm phosphat của nucleotide tiếp theo, tạo thành “khung xương sống” đường-phosphat luân phiên vững chắc.
Những base nitơ giữa hai mạch đơn polynucleotide link với nhau theo nguyên tắc bổ trợ ( A link với T, và C link với G ) trải qua những mối link hydro để tạo nên chuỗi DNA mạch kép. Tổng số lượng cặp base tương quan tới DNA trên Trái Đất ước tính bằng 5,0 x 1037, và nặng khoảng chừng 50 tỷ tấn. [ 5 ] Để so sánh, tổng khối lượng của sinh quyển xê dịch bằng 4 nghìn tỷ tấn carbon. [ 6 ]DNA tàng trữ thông tin sinh học, những mã di truyền đến những thế hệ tiếp theo và để hướng dẫn cho quy trình sinh tổng hợp protein. Mạch đơn DNA có link hóa học vững chãi chống lại sự phân cắt, và hai mạch đơn của chuỗi xoắn kép tàng trữ thông tin sinh học như nhau. tin tức này được sao chép nhờ sự phân tách hai mạch đơn. Một tỷ suất đáng kể DNA ( hơn 98 % ở người ) là những đoạn DNA không mã hóa ( non-coding ), nghĩa là những vùng này không giữ vai trò mạch khuôn để xác lập trình tự protein trải qua những quy trình phiên mã, dịch mã .
Hai mạch DNA chạy song song theo hai hướng ngược nhau. Gắn với mỗi phân tử đường là một trong bốn loại nucleobase (hay các base). Thông tin di truyền được mã hóa bởi trình tự của bốn nucleobase gắn trên mỗi mạch đơn. Những mạch RNA được tổng hợp từ những khuôn mẫu DNA trong quá trình phiên mã. Và dưới sự chỉ dẫn của mã di truyền, phân tử RNA tiếp tục được diễn dịch để xác định trình tự các amino acid ở cấu trúc protein trong quá trình dịch mã.
DNA ở tế bào nhân thực (động vật, thực vật, nấm và nguyên sinh vật) được lưu trữ bên trong nhân tế bào và một số bào quan, như ty thể hoặc lục lạp.[7] Ngược lại, ở sinh vật nhân sơ (vi khuẩn và vi khuẩn cổ), do không có nhân tế bào, DNA nằm trong tế bào chất. Bên trong tế bào, DNA tổ chức thành những cấu trúc dài gọi là nhiễm sắc thể (chromosome). Trong giai đoạn phân bào các nhiễm sắc thể hình thành được nhân đôi bằng cơ chế nhân đôi DNA, mang lại cho mỗi tế bào có một bộ nhiễm sắc thể hoàn chỉnh như nhau. Ở nhiễm sắc thể sinh vật nhân thực, những protein chất nhiễm sắc (chromatin) như histone giúp thắt chặt và tổ chức cấu trúc DNA. Chính cấu trúc thắt chặt này sẽ quản lý sự tương tác giữa DNA với các protein khác, quy định vùng nào của DNA sẽ được phiên mã.
Friedrich Miescher đã cô lập được DNA lần tiên phong vào năm 1869. Francis Crick và James Watson nhận ra cấu trúc phân tử chuỗi xoắn kép của nó vào năm 1953, dựa trên quy mô kiến thiết xây dựng từ tài liệu tích lũy qua ảnh chụp nhiễu xạ tia X do Rosalind Franklin triển khai. DNA trở thành một công cụ phân tử giúp những nhà nghiên cứu tò mò những kim chỉ nan và định luật vật lý sinh học, như định lý ergodic và triết lý đàn hồi. Những đặc thù vật tư độc lạ của DNA biến nó trở thành phân tử có ích so với những nhà khoa học vật tư chăm sóc trong nghành chế tạo vật liệu cỡ micro và nano, như trong công nghệ tiên tiến nano DNA. Các tân tiến trong nghành này gồm có chiêu thức origami DNA và vật tư lai dựa trên DNA. [ 8 ]
 Cấu trúc hóa học của DNA ; link hydro bộc lộ bằng những nét chấm .
Cấu trúc hóa học của DNA ; link hydro bộc lộ bằng những nét chấm . Cấu trúc phân tử 3 chiều của dạng thông dụng B-DNA .DNA là một polymer dài cấu trúc bởi những đơn phân nucleotide lặp lại. [ 9 ] [ 10 ] Cấu trúc DNA của mọi loài là không tĩnh ( non-static ), [ 11 ] chứa hai mạch polynucleotide xoắn đều quanh một trục tưởng tượng theo chiều từ trái sang phải ( xoắn phải ), mỗi một vòng xoắn ( khoảng chừng 10,4 cặp nucleotide ) dài 34 ångström ( 3,4 nm ) và có nửa đường kính 10 ångström ( 1,0 nm ). [ 12 ] Theo một điều tra và nghiên cứu khác, khi đo đạc trong những dung dịch đặc biệt quan trọng, chuỗi phân tử DNA rộng từ 22 đến 26 ångström ( 2,2 đến 2,6 nm ), và một đơn vị chức năng nucleotide dài 3,3 Å ( 0,33 nm ). [ 13 ] Dù cho mỗi đơn vị chức năng lặp lại có kích cỡ rất nhỏ, polymer DNA vẫn hoàn toàn có thể là những phân tử rất lớn chứa hàng triệu nucleotide. Ví dụ, DNA trong nhiễm sắc thể lớn nhất ở người, nhiễm sắc thể số 1, chứa xê dịch 220 triệu cặp base [ 14 ] và dài đến 85 mm nếu được duỗi thẳng .
Cấu trúc phân tử 3 chiều của dạng thông dụng B-DNA .DNA là một polymer dài cấu trúc bởi những đơn phân nucleotide lặp lại. [ 9 ] [ 10 ] Cấu trúc DNA của mọi loài là không tĩnh ( non-static ), [ 11 ] chứa hai mạch polynucleotide xoắn đều quanh một trục tưởng tượng theo chiều từ trái sang phải ( xoắn phải ), mỗi một vòng xoắn ( khoảng chừng 10,4 cặp nucleotide ) dài 34 ångström ( 3,4 nm ) và có nửa đường kính 10 ångström ( 1,0 nm ). [ 12 ] Theo một điều tra và nghiên cứu khác, khi đo đạc trong những dung dịch đặc biệt quan trọng, chuỗi phân tử DNA rộng từ 22 đến 26 ångström ( 2,2 đến 2,6 nm ), và một đơn vị chức năng nucleotide dài 3,3 Å ( 0,33 nm ). [ 13 ] Dù cho mỗi đơn vị chức năng lặp lại có kích cỡ rất nhỏ, polymer DNA vẫn hoàn toàn có thể là những phân tử rất lớn chứa hàng triệu nucleotide. Ví dụ, DNA trong nhiễm sắc thể lớn nhất ở người, nhiễm sắc thể số 1, chứa xê dịch 220 triệu cặp base [ 14 ] và dài đến 85 mm nếu được duỗi thẳng .
 Phân biệt cấu trúc nucleoside và nucleotide .
Phân biệt cấu trúc nucleoside và nucleotide .
Trong những sinh vật sống, DNA thường không tồn tại như một chuỗi đơn lẻ, mà thay vào đó là một cặp chuỗi liên kết chặt khít với nhau.[12][15] Hai mạch dài này quấn vào nhau như dây leo, tạo thành hình xoắn ốc kép. Một nucleobase liên kết với một phân tử đường tạo thành cấu trúc gọi là nucleoside, và một base liên kết với một phân tử đường và một hoặc nhiều nhóm phosphat gọi là nucleotide (nucleotide trong DNA và RNA là loại nucleotide chỉ mang một nhóm phosphat). Mạch polymer chứa nhiều nucleotide gắn kết với nhau (như trong DNA) được gọi là polynucleotide.[16] Mỗi nucleotide chứa cả hai thành phần đường và nhóm phosphat đóng vai trò khung xương cho phân tử (giữ cho các đơn phân của mạch liên kết với nhau), và chứa nucleobase để tương tác với mạch DNA còn lại trong chuỗi xoắn kép thông qua hệ thống liên kết hydro.
Khung xương chính của mạch DNA hình thành từ các nhóm phosphat và phân tử đường luân phiên nhau.[17] Phân tử đường trong DNA là 2-deoxyribose, một loại đường pentose (5 carbon). Các phân tử đường liên kết với nhau thông qua trung gian nhóm phosphat tạo thành liên kết phosphodieste giữa nguyên tử carbon thứ 3 với nguyên tử carbon thứ 5 trên hai mạch vòng của hai phân tử đường kế cận. Liên kết bất đối xứng này cho phép xác định hướng chạy của mạch đơn DNA. Xem xét gần hơn trên một chuỗi xoắn kép, người ta nhận thấy các nucleotide hướng theo một chiều trên một mạch và theo chiều ngược lại trên mạch kia, gọi là: hai mạch hướng ngược chiều nhau hay đối song song (antiparallel). Các đầu không đối xứng kết thúc của chuỗi DNA là đầu 5′ (năm phẩy) và đầu 3′ (ba phẩy), với đầu 5′ kết thúc bởi nhóm phosphat và đầu 3′ kết thúc bởi nhóm hydroxyl (OH). Sự khác nhau chủ yếu giữa DNA và RNA là ở phân tử đường, với đường 2-deoxyribose trong DNA được thay thế bởi đường ribose trong RNA.[15]
Hai mạch xoắn của chuỗi DNA được gắn không thay đổi bởi hai lực link chính : link hydro giữa những nucleotide của hai mạch và tương tác chồng chất ( base-stacking ) giữa những base thơm. [ 19 ] Trong môi trường tự nhiên dung dịch của tế bào, link π phối hợp của những base nucleotide sắp xếp vuông góc với trục của phân tử DNA, giảm thiểu tương tác của chúng với lớp vỏ solvat hóa ( solvation shell ), và do vậy làm giảm nguồn năng lượng tự do Gibbs. Bốn base trong DNA là adenine ( viết tắt A ), cytosine ( C, ở Nước Ta còn viết là xitôzin, viết tắt X ), guanine ( G ) và thymine ( T ). Bốn base này gắn với nhóm đường / phosphat để tạo thành nucleotide hoàn hảo, như adenosine monophosphate. Adenine ghép cặp với thymine và guanine ghép cặp với cytosine, ký hiệu bằng những cặp base A-T và G-C. [ 20 ] [ 21 ]
Mục lục nội dung
Phân loại nucleobase[sửa|sửa mã nguồn]
Các nucleobase được phân thành hai loại : purine, gồm adenine ( A ) và guanine ( G ), là hợp chất dị vòng có hai vòng 5 và 6 nguyên tử carbon gắn với nhau ; và pyrimidine, gồm cytosine ( C ) và thymine ( T ), là hợp chất dị vòng có 6 nguyên tử carbon. [ 15 ] Một nucleobase pyrimidine thứ năm là uracil ( U ), thay thế sửa chữa cho thymine ( T ) trong RNA và khác với thymine do thiếu đi một nhóm methyl ( – CH3 ) trên vòng của nó. Ngoài RNA và DNA, một số lượng lớn acid nucleic tự tạo tựa như được tạo ra để nghiên cứu và điều tra những đặc thù của acid nucleic, hoặc sử dụng trong công nghệ sinh học. [ 22 ]
Uracil thường không có ở DNA, nó chỉ xuất hiện như một sản phẩm phân tách của cytosine. Tuy nhiên, ở một số thực khuẩn thể như: thực khuẩn thể ở Bacillus subtilis PBS1 và PBS2 và thực khuẩn Yersinia piR1-37, thì thymine được thay bằng uracil.[23] Một thực khuẩn thể khác – thể Staphylococcal S6 – được phát hiện với bộ gene mà thymine thay bằng uracil.[24]
Base J (beta-d-glucopyranosyloxymethyluracil), một dạng tinh chỉnh của uracil, cũng xuất hiện ở một số sinh vật: trùng roi Diplonema và Euglena, và mọi nhóm Kinetoplastida.[25] Sinh tổng hợp base J diễn ra theo hai bước: bước thứ nhất một thymidine xác định trong DNA được biến đổi thành hydroxymethyldeoxyuridine (HOMedU); bước thứ hai HOMedU được glycosyl hóa thành base J.[26] Các nhà khoa học cũng khám phá ra những protein được tổng hợp từ base này.[27][28][29] Những protein này dường như có họ hàng xa với gene gây ung thư (oncogene) Tet1 mà tham gia vào quá trình phát sinh bệnh bạch cầu myeloid cấp tính.[30] Base J cũng đóng vai trò làm tín hiệu kết thúc cho enzyme RNA polymerase II.[31][32]
 Rãnh lớn và rãnh nhỏ trên phân tử DNA. Rãnh nhỏ là một vị trí link với chất nhuộm màu Hoechst 33258
Rãnh lớn và rãnh nhỏ trên phân tử DNA. Rãnh nhỏ là một vị trí link với chất nhuộm màu Hoechst 33258
Hai mạch đơn xoắn đôi vào nhau tạo thành bộ khung cho DNA. Ở chuỗi xoắn kép này có thể xuất hiện những khoảng trống nằm cách nhau giữa hai mạch gọi là các rãnh (groove). Những rãnh này nằm liền kề với các cặp base và có thể hình thành một điểm bám (binding site). Vì hai mạch đơn không đối xứng nhau nên dẫn đến các rãnh có kích thước không đều, trong đó rãnh lớn (major groove) rộng 22 Å và rãnh nhỏ (minor groove) rộng 12 Å.[33] Độ rộng của rãnh giúp cho các cạnh của base trở nên dễ tiếp cận hơn trong rãnh lớn so với rãnh nhỏ. Kết quả là, các protein của các nhân tố phiên mã mà liên kết với những đoạn trình tự cụ thể trong chuỗi xoắn kép DNA thường thực hiện bằng việc tiếp xúc với các cạnh của các base ở rãnh lớn.[34] Tình huống này thay đổi đa dạng tùy theo hình dáng bất thường của DNA bên trong tế bào (xem ở dưới), nhưng các rãnh lớn và rãnh nhỏ luôn luôn được đặt tên để phản ánh sự khác nhau về kích thước đo được nếu DNA vặn xoắn trở về dạng B thường gặp.
Trong chuỗi xoắn kép DNA, mỗi loại nucleobase trên một mạch chỉ link với một loại nucleobase trên mạch kia. Đây được gọi là nguyên tắc bổ trợ cặp base. Ở đây, purine hình thành link hydro với pyrimidine, trong đó adenine chỉ ghép với thymine bằng hai link hydro, và cytosine chỉ ghép với guanine bằng ba link hydro. Sự sắp xếp giữa hai nucleotide link với nhau qua chuỗi xoắn kép gọi là một cặp base. Vì link hydro không phải là link cộng hóa trị, nên hoàn toàn có thể bị đứt ra và nối lại tương đối thuận tiện. Hai mạch của DNA trong chuỗi xoắn kép do vậy hoàn toàn có thể tách rời nhau ra giống như khóa kéo, hoặc bằng lực cơ học hoặc bằng nhiệt độ cao. [ 35 ] Hệ quả của nguyên tắc bổ trợ này là mọi thông tin trong trình tự chuỗi xoắn kép DNA được lặp lại ở mỗi mạch, và có vai trò quan trọng trong tiến trình sao chép DNA. Nói chung, trình tự lặp lại ngược chiều giữa hai mạch và những tương tác link bổ trợ trong những cặp base là tối quan trọng so với mọi công dụng của DNA trong khung hình sống. [ 10 ]


Hình trên, cặp base G-C liên kết bằng ba liên kết hydro. Hình dưới, cặp base A-T liên kết bằng hai liên kết hydro. Liên kết hydro không phải là liên kết cộng hóa trị và được thể hiện bằng các nét chấm nhỏ.
Hai loại cặp base khác nhau bởi số link hydro giữa những base, cặp A-T có 2 link hydro và cặp G-C có 3 link hydro. Những phân tử DNA chứa nhiều cặp G-C sẽ không thay đổi hơn so với những phân tử chứa ít cặp G-C .
Như miêu tả ở trên, hầu hết phân tử DNA bao gồm hai mạch polymer liên kết thành dạng xoắn kép bởi liên kết hydro không phải là liên kết cộng hóa trị; cấu trúc mạch kép này (dsDNA – double stranded DNA) cũng được duy trì chủ yếu bởi những tương tác chồng chất pi giữa các base trên hai mạch, mà mạnh nhất là ở cấu trúc chồng chất G,C (tương tác chồng chất pi là những tương tác không cộng hóa trị giữa các vòng thơm mang liên kết pi liên hợp). Hai mạch có thể tách nhau ra – một quá trình gọi là nóng chảy – để tạo thành hai phân tử DNA mạch đơn (ssDNA – single-stranded DNA). Sự phân tách xảy ra ở nhiệt độ cao, độ mặn thấp và độ pH cao (độ pH thấp cũng làm tách DNA, nhưng vì DNA trở nên không ổn định do acid bị khử purine hóa (bản chất DNA là một loại acid), do đó độ pH thấp ít khi được sử dụng).
Sự không thay đổi của dạng mạch kép dsDNA không riêng gì nhờ vào vào thành phần G-C ( tỷ suất % cặp base G-C ) mà còn phụ thuộc vào vào trình tự những base ( do tương tác chồng chất pi giữa những base là một thuộc tính đặc hiệu của trình tự ) và độ dài ( phân tử càng dài thì càng không thay đổi ). Độ không thay đổi được đo bằng nhiều cách khác nhau ; cách phổ cập là đưa phân tử đạt tới ” nhiệt độ nóng chảy “, đó là nhiệt độ mà tại đấy khoảng chừng 50 % số phân tử ds đổi khác thành phân tử ss ; nhiệt độ nóng chảy phụ thuộc vào vào cường độ ion và sự đông đặc của DNA. Do vậy, cả tỷ suất Xác Suất số cặp base G-C và chiều dài toàn diện và tổng thể của chuỗi xoắn kép DNA xác lập nên cường độ link giữa hai mạch DNA. Những chuỗi xoắn kép DNA dài với thành phần nhiều G-C có tương tác giữa hai mạch mạnh hơn so với những chuỗi xoắn kép ngắn với thành phần nhiều A-T. [ 36 ] Trong hoạt động giải trí sinh học, có những phần của chuỗi xoắn kép DNA thuận tiện tách ra khi thiết yếu, ví dụ như hộp Pribnow TATAAT ở một số ít vùng khởi động ( promoter ), có khuynh hướng chứa nhiều thành phần A-T, khiến cho những mạch hoàn toàn có thể phân tách thuận tiện. [ 37 ]
Trong phòng thí nghiệm, cường độ của tương tác này có thể đo bằng cách tìm ra nhiệt độ cần thiết để phân cắt liên kết hydro giữa hai mạch, hay chính là nhiệt độ nóng chảy của chúng (được ký hiệu là Tm, nghĩa là melting temperature (nhiệt độ nóng chảy)). Khi tất cả các cặp base tách rời nhau, hai mạch của chuỗi DNA sẽ tách rời và tồn tại trong dung dịch như các phân tử độc lập. Những phân tử mạch đơn DNA (ssDNA) không có hình dạng chung, nhưng một số có thể thu về những dạng ổn định tùy theo độ dài và thành phần cặp base.[38]
Có nghĩa và đối nghĩa[sửa|sửa mã nguồn]
Một trình tự DNA gọi là “có nghĩa” (sense) nếu trình tự của nó giống với trình tự của bản sao RNA thông tin dùng để dịch mã thành protein.[39] Khi đó, trình tự trên mạch bổ sung còn lại được gọi là trình tự “đối nghĩa” (antisense). Cả trình tự có nghĩa và đối nghĩa có thể tồn tại trên các đoạn khác nhau của cùng một mạch đơn DNA (tức là cả hai mạch có thể chứa cả trình tự có nghĩa lẫn đối nghĩa). Ở tế bào nhân thực và nhân sơ, các trình tự RNA đối nghĩa đều được tạo ra, nhưng chức năng của những RNA này vẫn chưa được hiểu rõ hoàn toàn.[40] Có đề xuất cho rằng các RNA đối nghĩa có khả năng tham gia vào hoạt động điều hòa biểu hiện gene thông qua sự bổ sung base RNA-RNA.[41]
Một vài trình tự DNA ở sinh vật nhân thực và nhân sơ, và hay gặp hơn ở plasmid và virus, xóa nhòa sự độc lạ giữa những mạch có nghĩa và đối nghĩa do có sự hiện hữu của những gene chồng lợp ( overlapping gene ). [ 42 ] Trong trường hợp này, 1 số ít trình tự DNA tiếp đón đến hai nghĩa vụ và trách nhiệm, mã hóa cho một protein khi đọc dọc theo một mạch, và mã hóa protein thứ hai khi đọc theo hướng ngược lại dọc theo mạch kia. Trong vi trùng, sự chồng lợp này hoàn toàn có thể ảnh hưởng tác động đến quy trình điều hòa phiên mã gene, [ 43 ] trong khi ở virus, những gene chồng lợp lại làm tăng lượng thông tin được mã hóa bên trong bộ gene nhỏ bé của virus. [ 44 ]
DNA siêu xoắn[sửa|sửa mã nguồn]
DNA có thể xoắn lại tựa như một sợi dây thừng theo một tiến trình gọi là DNA siêu xoắn (DNA supercoiling). Với DNA ở trạng thái “bình thường”, một mạch thường xoắn đều quanh trục tưởng tượng của chuỗi xoắn kép theo từng đoạn ngắn mang khoảng 10,4 cặp base, nhưng nếu DNA bị vặn xoắn thì các mạch có thể trở nên siết chặt hơn hoặc lỏng lẻo hơn.[45] Nếu DNA bị xoắn theo hướng của chuỗi xoắn kép, hay siêu xoắn thuận (positive supercoiling), thì các base giữ chặt với nhau hơn. Còn nếu DNA bị xoắn ngược hướng với chuỗi xoắn kép, hay siêu xoắn nghịch (negative supercoiling), thì các base phân tách dễ dàng hơn. Trong tự nhiên, hầu hết DNA trong tế bào đều ở trạng thái gần siêu xoắn nghịch do chịu sự tác động của nhóm enzyme có tên gọi topoisomerase.[46] Những enzyme này cũng cần thiết để tháo xoắn các mạch DNA trong những quá trình như phiên mã và nhân đôi DNA.[47]
 Từ trái qua phải, những cấu trúc của DNA dạng A, B và Z .
Từ trái qua phải, những cấu trúc của DNA dạng A, B và Z .
Những quy mô cấu trúc DNA[sửa|sửa mã nguồn]
DNA hoàn toàn có thể sống sót ở nhiều thông số kỹ thuật, trong đó gồm có A-DNA, B-DNA, và Z-DNA, mặc dầu chỉ có thông số kỹ thuật B-DNA và Z-DNA trực tiếp quan sát thấy trong những sinh vật chuyên hóa công dụng. [ 17 ] Cấu hình mà DNA tuân theo nhờ vào vào mức độ hydrat hóa, trình tự DNA, số cặp base và khunh hướng siêu xoắn, cộng với những tu sửa hóa học trên những base, thành phần và hàm lượng ion sắt kẽm kim loại, cũng như sự hiện hữu của những polyamin trong dung dịch. [ 48 ]
Báo cáo đầu tiên về ảnh chụp tán xạ tia X của dạng A-DNA và B-DNA sử dụng phương pháp phân tích dựa trên hàm Patterson chỉ cung cấp thông tin giới hạn về cấu trúc của các sợi định hướng trong DNA.[49][50] Một hướng phân tích khác, do Wilkins cùng cộng sự (et al.) đề xuất vào năm 1953, cho các phần chụp nhiễu xạ-tán xạ tia X đối với B-DNA in vivo (trong cơ thể sống thí nghiệm) của các sợi DNA hydrat hóa cao độ tuân theo những hạng tử bình phương trong hàm Bessel.[51] Trong cùng tạp chí, James Watson và Francis Crick trình bày mô hình phân tử DNA của họ sau khi phân tích các hình ảnh nhiễu xạ tia X và gợi ra rằng cấu trúc của nó có dạng chuỗi xoắn kép.[12]
Dạng B-DNA là cấu hình phổ biến nhất tìm thấy dưới những điều kiện của tế bào sống,[52] tồn tại ở trạng thái gần giống tinh thể (paracrystalline state), đó là cấu hình động mặc dù tính tương đối cứng của chuỗi xoắn kép DNA được giữ ổn định bởi liên kết hydro giữa các base. Để đơn giản, hầu hết những mô hình phân tử DNA đều bỏ qua liên kết động lực của nước và các ion đối với phân tử dạng B-DNA, và do đó ít hữu ích khi dùng các mô hình này để hiểu cách hoạt động của B-DNA trong tế bào sống ở trạng thái bình thường (in vivo).[53] Phân tích vật lý và toán học của ảnh chụp tia X[54][55] cũng như dữ liệu quang phổ thu được cho dạng B-DNA tiền tinh thể (paracrystalline), do vậy phức tạp hơn so với dữ liệu nhiễu xạ tia X của ảnh chụp dạng A-DNA.
So với B-DNA, dạng A-DNA xoắn ốc theo chiều tay phải rộng hơn về đường kính, với một rãnh nhỏ nông hơn và rộng hơn, trong khi rãnh lớn sâu hơn và hẹp hơn. Dạng A thường Open dưới những điều kiện kèm theo phi sinh lý, đặc biệt quan trọng trong những mẫu DNA mất nước một phần, trong khi ở tế bào nó hoàn toàn có thể ở dạng lai ghép mạch đơn DNA với mạch đơn RNA, cũng như Open cả ở phức hệ enzyme-DNA. [ 56 ] [ 57 ] Ở đoạn DNA nơi những base đã bị tinh sửa về mặt hóa học bằng giải pháp methyl hóa hoàn toàn có thể trải qua sự đổi khác lớn về hình dạng thông số kỹ thuật và trở thành dạng Z-DNA. Ở thông số kỹ thuật đây, hai mạch xoắn quanh trục theo chiều tay trái, ngược chiều với hướng của dạng B thông dụng. [ 58 ] Những cấu trúc không bình thường này hoàn toàn có thể nhận ra bằng một loại protein đặc hiệu link với Z-DNA và những protein này hoàn toàn có thể tham gia vào hoạt động giải trí điều hòa quy trình phiên mã. [ 59 ]
Đặc điểm cấu trúc của ba cấu hình chính DNA[60][61][62]
Ghi chú: bp là cặp base (base pair)
Đặc tính hình học
A-DNA
B-DNA
Z-DNA
Chiều xoắn
phải
phải
trái
Đường kính
≈ 2,3 nm
≈ 2,0 nm
≈ 1,8 nm
Đơn vị lặp lại
1 bp
1 bp
2 bp
Góc quay/bp
32,7°
34,3°
60°/2
Số bp trung bình/vòng xoắn
11
10,4
12
Độ nghiêng của bp so với trục
+19°
-1,2°
-9°
Độ dài dốc/bp dọc theo trục
0,23 nm
0,332 nm
0,38 nm
Bước/vòng xoắn
2,82 nm
3,32 nm
4,56 nm
Góc xoắn trung bình giữa hai bp
+18°
+16°
0°
Góc glycosyl
anti
anti
Pyrimidine: anti
Purine: syn
Chế độ gấp phân tử đường
(sugar puckering)
C3′-endo
C2′-endo
Pyrimidine: C2′-endo
Purine: C3′-endo
Rãnh lớn
hẹp và sâu
rộng và sâu, độ sâu: 0,85 nm
phẳng
Rãnh nhỏ
rộng và phẳng
hẹp và sâu, độ sâu: 0,75 nm
hẹp và sâu
DNA có thành phần hóa học thay thế sửa chữa[sửa|sửa mã nguồn]
Trong một vài năm, những nhà sinh học thiên hà đã yêu cầu về một sinh quyển bóng tối ( shadow biosphere ), một sinh quyển vi sinh vật giả thuyết sống sót trên Trái Đất mà sử dụng những quy trình phân tử và hóa học khác cơ bản so với những gì đã biết về sự sống hiện tại. Một trong những đề xuất kiến nghị đó là sự sống sót của dạng sinh vật sống mà nguyên tử asen thay cho phospho trong DNA. Một báo cáo giải trình năm 2010 cho thấy năng lực này xuất hiện trong vi trùng GFAJ-1, [ 63 ] [ 63 ] [ 64 ] mặc dầu đã có những tranh cãi, [ 64 ] [ 65 ] và ở đầu cuối năm 2012 một báo cáo giải trình khác nêu ra vật chứng cho thấy những vi trùng này dữ thế chủ động ngăn không cho asen phối hợp vào bộ khung DNA của nó và những phân tử sinh học khác. [ 66 ]
Cấu trúc bộ bốn[sửa|sửa mã nguồn]
Xem thêm thông tin : G-quadruplexTại đầu mút của mỗi nhiễm sắc thể là những vùng đặc hiệu của DNA gọi là telomere. Chức năng chính của nhóm vùng này đó là được cho phép tế bào thực thi sao chép những đầu mút nhiễm sắc thể sử dụng enzyme telomerase, chính do thông thường những enzyme sao chép DNA không thể nhân đôi đến đầu 3 ′ tận cùng của nhiễm sắc thể. [ 67 ] Những đầu mút đặc hiệu này của nhiễm sắc thể cũng giúp bảo vệ DNA bị rút ngắn sau mỗi lần nhân đôi, và cho dừng mạng lưới hệ thống thay thế sửa chữa DNA trong tế bào khi mạng lưới hệ thống này coi DNA bị hỏng và cần được sửa chữa thay thế. [ 68 ] Trong tế bào người, những telomere thường là những mạch đơn DNA dài chứa vài nghìn trình tự TTAGGG lặp đi lặp lại. [ 69 ]
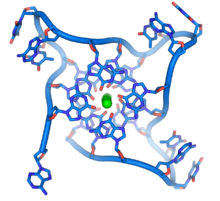 [70]Cấu trúc bộ bốn DNA hình thành bằng những đoạn telomere lặp lại. Hình dạng vòng của bộ khung DNA nhìn rất khác so với dạng xoắn ốc của DNA nổi bật. Những hình cầu xanh lục ở giữa đại diện thay mặt cho những ion kali .
[70]Cấu trúc bộ bốn DNA hình thành bằng những đoạn telomere lặp lại. Hình dạng vòng của bộ khung DNA nhìn rất khác so với dạng xoắn ốc của DNA nổi bật. Những hình cầu xanh lục ở giữa đại diện thay mặt cho những ion kali .
Các trình tự giàu guanine có khả năng giữ ổn định những đầu mút của nhiễm sắc thể bằng cách hình thành nên cấu trúc gồm những đơn vị chứa bốn base xếp chồng lên nhau, hơn là dạng bổ sung cặp base thường thấy ở các phân tử DNA khác. Ở đây, bốn base guanine tạo thành một tấm phẳng và những đơn vị phẳng chứa bốn base này xếp xen chồng lẫn nhau hình thành nên cấu trúc G-quadruplex (bộ bốn) ổn định.[71] Sự ổn định của cấu trúc này có được là do liên kết hydro giữa các cạnh của base và hiện tượng chelat hóa của một ion kim loại nằm ở trung tâm của khối phẳng bộ bốn base.[72] Những cấu trúc khác cũng có thể tồn tại, với trung tâm của bộ bốn base hoặc là một mạch đơn gấp xoắn xung quanh các base, hoặc là một vài mạch song song với nhau, trong đó mỗi mạch đều đóng góp một base vào cấu trúc trung tâm.
Bên cạnh dạng cấu trúc xếp chồng, telomere cũng có cấu trúc dạng vòng lớn gọi là vòng telomere (telomere loop), hay T-loop. Trong cấu trúc này, một mạch đơn DNA quấn quanh thành một vòng tròn dài ổn định bởi các protein liên kết với telomere.[73] Tại đầu mút tận cùng của T-loop, telomere mạch đơn DNA được giữ ở một vùng bao bởi DNA mạch kép bằng mạch telomere phân tách mạch kép DNA và thực hiện việc bổ sung cặp base với một trong hai mạch. Cấu trúc ba mạch này (triple-stranded DNA) được gọi là vòng chuyển chỗ (displacement loop) hay D-loop.[71]
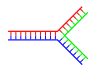

Nhánh đơn mạch
Nhánh đa mạch
DNA phân nhánh hoàn toàn có thể tạo thành mạng lưới chứa nhiều nhánh .
DNA phân nhánh[sửa|sửa mã nguồn]
Ở chuỗi xoắn kép DNA, hiện tượng kỳ lạ sờn tước đầu mút Open khi những đoạn không được bổ trợ hiện hữu tại đầu mút của DNA mạch kép. Qua đó, DNA phân nhánh hoàn toàn có thể hình thành nếu có một mạch DNA thứ ba Open và mang những đoạn mới phối hợp với đoạn không được bổ trợ của chuỗi xoắn kép đã bị sờn tước trước đó. Dạng đơn thuần nhất của DNA phân nhánh chỉ gồm có ba mạch DNA, tất yếu là hoàn toàn có thể sống sót thêm nhiều nhánh phức tạp khác. [ 74 ] DNA phân nhánh được ứng dụng trong công nghệ tiên tiến nano để lắp ráp những thông số kỹ thuật phân tử mong ước .
Những biến hóa hóa học và trình tự của DNA[sửa|sửa mã nguồn]
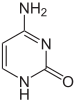


cytosine
5-methylcytosine
thymine
Cấu trúc của cytosine khi có và không có nhóm 5 – methyl. Sự khử amin biến hóa 5 – methylcytosine thành thymine .
Chỉnh sửa base và phương cách đóng gói DNA[sửa|sửa mã nguồn]
Biểu hiện của gene chịu ảnh hưởng bởi phương cách đóng gói DNA trong nhiễm sắc thể, thành một cấu trúc gọi là chất nhiễm sắc (chromatin). Những tác động chỉnh sửa base có thể xảy ra trong quá trình đóng gói, với các vùng không có hoặc có mức biểu hiện gene thấp thông thường chứa các base cytosine ở mức methyl hóa cao. Sự đóng gói DNA và ảnh hưởng của nó lên biểu hiện gene cũng xảy ra bởi hiệu ứng thay đổi liên kết cộng hóa trị tại lõi protein histone bọc quanh DNA trong cấu trúc chất nhiễm sắc hoặc bởi phức hệ chất nhiễm sắc tái mô hình hóa (xem Tái mô hình hóa chất nhiễm sắc (chromatin remodeling)). Do vậy, tác động xen lẫn giữa methyl hóa DNA và thay đổi liên kết ở histone có ảnh hưởng phối hợp đến chất nhiễm sắc và biểu hiện gene.[75]
Ví dụ, sự methyl hóa cytosine, tạo ra 5-methylcytosine, có vai trò quan trọng đối với sự bất hoạt X của nhiễm sắc thể (X-inactivation).[76] Mức độ methyl hóa trung bình thay đổi theo mỗi sinh vật – giun tròn Caenorhabditis elegans không có phản ứng methyl hóa cytosine, trong khi ở động vật có xương sống có mức độ cao hơn, lên tới 1% lượng DNA chứa 5-methylcytosine.[77] Tuy 5-methylcytosine có vai trò quan trọng, nhưng nó vẫn có thể bị khử amin hóa để chuyển thành base thymine, do đó cytosine methyl hóa có khuynh hướng gây đột biến.[78] Những thay đổi base khác bao gồm sự methyl hóa adenine ở vi khuẩn, sự hiện diện của 5-hydroxymethylcytosine trong não,[79] và sự glycosyl hóa của uracil tạo thành “Base J” trong các loài Kinetoplastida.[80][81]
DNA hoàn toàn có thể bị hư hại bởi nhiều tác nhân đột biến, làm đổi khác trình tự DNA. Những tác nhân đột biến gồm có những chất oxy hóa, những chất ankyl hóa cũng như bức xạ điện từ nguồn năng lượng cao như tia cực tím và tia X. Loại DNA hư hại hình thành phụ thuộc vào vào loại tác nhân đột biến. Ví dụ, tia UV hoàn toàn có thể tàn phá DNA khi tạo ra thymine nhị trùng ( thymine dimer ), nghĩa là cấu thành link chéo giữa những base pyrimidine với nhau. [ 83 ] Mặt khác, những tác nhân oxy hóa như gốc tự do hay hydro peroxid tạo ra nhiều dạng hư hại, gồm có tinh sửa base, đặc biệt quan trọng là guanosine, và làm đứt gãy chuỗi xoắn kép. [ 84 ] Một tế bào nổi bật ở người chứa khoảng chừng 150.000 base chịu sự hủy hoại dưới tác nhân oxy hóa. [ 85 ] Trong những tổn hại oxy hóa này, mức độ nguy khốn nhất đó là làm chuỗi xoắn kép bị đứt gãy, vì rất khó để hàn gắn chúng lại và hoàn toàn có thể dẫn tới đột biến điểm ( point mutation ), đột biến thêm đoạn và mất đoạn trên trình tự DNA, cũng như quy trình chuyển đoạn nhiễm sắc thể ( chromosomal translocation ). [ 86 ] Những đột biến này hoàn toàn có thể gây ra ung thư. Bởi vì những số lượng giới hạn vốn có trong chính sách thay thế sửa chữa DNA, nếu con người sống đủ lâu, những hư hại này ở đầu cuối sẽ dẫn tới sự tăng trưởng của ung thư. [ 87 ] [ 88 ] Những tàn phá DNA mà Open một cách tự nhiên là do những quy trình thông thường trong tế bào tạo ra những mẫu sản phẩm phản ứng với oxy, ví dụ điển hình những phản ứng thủy phân của nước trong tế bào, v.v, cũng xảy ra một cách tiếp tục. Mặc dù hầu hết những tàn phá này đều được sửa chữa thay thế, nhưng trong bất kể tế bào nào vẫn có một vài DNA hư hại hoàn toàn có thể còn sống sót mặc cho những hoạt động giải trí sửa chữa thay thế. Những DNA bị hủy hoại còn sót lại sẽ tích tụ dần theo độ tuổi bên trong những mô sau nguyên phân ở động vật hoang dã. Sự tích tụ này có vẻ như là một nguyên do quan trọng dẫn tới sự già yếu. [ 89 ] [ 90 ] [ 91 ]
Nhiều tác nhân đột biến nằm gọn trong không gian giữa hai cặp base liền kề, hay gọi là các phân tử xen kẹp (intercalation). Hầu hết các phân tử xen kẹp là những hợp chất vòng thơm cấu trúc phẳng; ví dụ: ethidium bromide, acridine, daunomycin và doxorubicin. Để cho một phân tử xen kẹp có thể vừa vặn không gian giữa hai cặp base, các base phải bị tách ra, bóp méo chuỗi DNA bằng cách tháo xoắn mạch kép. Điều này ngăn cản quá trình phiên mã và nhân đôi DNA, phát xuất độc tính và những đột biến.[92] Kết quả là, các phân tử xen kẹp vào DNA có thể là tác nhân gây ung thư, và trong trường hợp của thalidomide là tác nhân gây quái thai (teratogen).[93] Những phân tử khác như benzo[a]pyrene diol epoxide và aflatoxin tạo thành sản phẩm cộng vào DNA dẫn tới các lỗi trong quá trình nhân đôi.[94] Tuy thế, do khả năng ngăn cản sự phiên mã và nhân đôi DNA, những độc tố tương tự khác cũng được sử dụng trong phương pháp hóa trị liệu để ngăn chặn sự lớn lên nhanh chóng của các tế bào ung thư.[95]
Chức năng sinh học[sửa|sửa mã nguồn]
 Vị trí của DNA nhân chứa trong chất nhiễm sắc bên trong nhân tế bào của tế bào nhân thực .
Vị trí của DNA nhân chứa trong chất nhiễm sắc bên trong nhân tế bào của tế bào nhân thực .
DNA thông thường hiện diện trong nhiễm sắc thể dạng thẳng ở sinh vật nhân thực, và nhiễm sắc thể dạng vòng ở sinh vật nhân sơ. Nhiễm sắc thể (chromosome) thực chất là chất nhiễm sắc (chromatin) bị co xoắn từ kỳ đầu của quá trình phân bào. Còn chất nhiễm sắc chính là phức hợp giữa chuỗi xoắn kép DNA với các protein histone và phi histone gói gọn thành một cấu trúc cô đặc. Điều này cho phép các phân tử DNA rất dài nằm gọn trong nhân tế bào. Cấu trúc vật lý của nhiễm sắc thể và chất nhiễm sắc thay đổi luân phiên tùy thuộc vào từng giai đoạn của chu kỳ tế bào. Tập hợp các nhiễm sắc thể trong một tế bào tạo thành bộ gene của nó; bộ gene người có xấp xỉ 3 tỷ cặp base DNA xếp thành 46 nhiễm sắc thể.[96] Thông tin chứa trong DNA tổ chức dưới dạng trình tự của các đoạn DNA gọi là gene. Sự kế thừa thông tin di truyền trong gene được thực hiện thông qua các cặp base bổ sung. Ví dụ, trong quá trình phiên mã, khi một tế bào sử dụng thông tin ở một gene, trình tự DNA sẽ được sao mã vào trình tự bổ sung RNA thông qua lực hút giữa DNA và các nucleotide chính xác của RNA. Thông thường, bản sao RNA này được dùng làm khuôn mẫu để xác định trình tự các amino acid trong quá trình dịch mã, thông qua sự tương tác giữa các nucleotide RNA. Trong quá trình khác, một tế bào có thể tự sao chép thông tin di truyền của nó bằng quá trình nhân đôi DNA. Chi tiết của những chức năng này được nêu trong những bài viết liên quan; bài này tập trung vào tương tác giữa DNA và các phân tử khác mà đảm trách các chức năng của bộ gene.
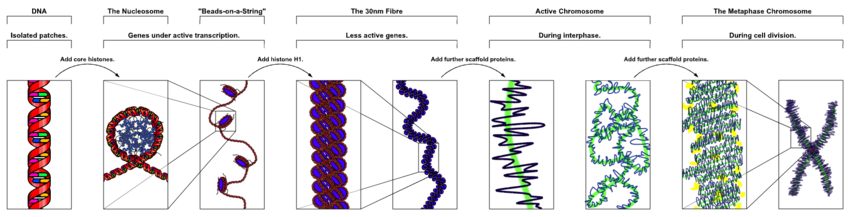 Hình minh họa những mức độ co xoắn từ DNA đến nhiễm sắc thể kép tại kỳ giữa của quy trình phân bào : Chuỗi xoắn kép DNA 2 nm, cấu trúc nucleosome, chuỗi nucleosome ( sợi cơ bản ) 10 nm, sợi chất nhiễm sắc 30 nm, sợi siêu xoắn 300 nm, chromatid 700 nm và nhiễm sắc thể kép 1400 nm ở mức xoắn cực lớn. Xem video trực quan
Hình minh họa những mức độ co xoắn từ DNA đến nhiễm sắc thể kép tại kỳ giữa của quy trình phân bào : Chuỗi xoắn kép DNA 2 nm, cấu trúc nucleosome, chuỗi nucleosome ( sợi cơ bản ) 10 nm, sợi chất nhiễm sắc 30 nm, sợi siêu xoắn 300 nm, chromatid 700 nm và nhiễm sắc thể kép 1400 nm ở mức xoắn cực lớn. Xem video trực quan
Gene và bộ gene[sửa|sửa mã nguồn]
 Gene là một đoạn của DNA mã hóa những thông tin công dụng sinh học. Nhiễm sắc thể chứa một chuỗi DNA dài trên đó gồm có rất nhiều gene. Một nhiễm sắc thể ở người hoàn toàn có thể chứa tới 500 triệu cặp base với hàng nghìn gene .
Gene là một đoạn của DNA mã hóa những thông tin công dụng sinh học. Nhiễm sắc thể chứa một chuỗi DNA dài trên đó gồm có rất nhiều gene. Một nhiễm sắc thể ở người hoàn toàn có thể chứa tới 500 triệu cặp base với hàng nghìn gene .
DNA chứa các đoạn gene được gói gọn và xếp chặt có thứ tự bởi quá trình cô đặc DNA (DNA condensation), để có thể vừa vặn trong một thể tích nhỏ của tế bào. Ở sinh vật nhân thực, DNA nằm trong nhân tế bào, cùng với một lượng nhỏ nằm trong ty thể và lục lạp. Ở sinh vật nhân sơ, DNA nằm trong một thể có hình dạng không đều giữa tế bào chất, gọi là thể nhân (hoặc vùng nhân, nucleoid).[97] Thông tin di duyền trong một bộ gene được lưu trữ bởi các gene, và tập hợp toàn bộ các gene trong tế bào của cơ thể thuộc một loài sinh vật được gọi là kiểu gene. Mỗi gene là một đơn vị của tính di truyền và là một đoạn của DNA có ảnh hưởng tới một đặc tính cụ thể trong cơ thể sinh vật. Các gene chứa một khung đọc mở (open reading frame) có thể được phiên mã, cùng với các vùng trình tự điều hòa (regulatory sequence) như vùng khởi động (promoter) và vùng tăng cường (enhancer) có khả năng điều hòa quá trình phiên mã của khung đọc mở.
Ở nhiều loài, chỉ một phần nhỏ trong tổng số trình tự của bộ gene là mã hóa cho protein. Ví dụ, chỉ khoảng 1,5% bộ gene người chứa các đoạn exon mã hóa cho protein, trong khi trên 50% DNA ở người chứa các trình tự lặp lại không mã hóa (non-coding repeated sequence).[98] Những lý do cho sự có mặt của rất nhiều DNA không mã hóa ở bộ gene của sinh vật nhân thực và sự cách biệt rất lớn trong kích cỡ bộ gene, hay giá trị C, giữa các loài đã đưa đến một vấn đề nan giải lâu năm gọi là “nghịch lý giá trị C”.[99] Tuy nhiên, một số trình tự DNA không mã hóa protein vẫn có thể có chức năng mã hóa các phân tử RNA không mã hóa tham gia vào quá trình điều hòa biểu hiện gene.[100]
Xem thêm: LGBT – Wikipedia tiếng Việt
Một số trình tự DNA không mã hóa đóng vai trò cấu trúc bộ khung trong nhiễm sắc thể. Telomere và tâm động ( centromere ) nổi bật chỉ chứa vài gene, nhưng lại có vai trò quan trọng so với công dụng và sự không thay đổi của nhiễm sắc thể. [ 68 ] [ 102 ] Một dạng DNA không mã hóa Open ở người gọi là gene giả ( pseudogene ), là những bản sao của gene nhưng đã bị bất hoạt do tác động ảnh hưởng của đột biến. [ 103 ] Những trình tự này thường chỉ là những hóa thạch phân tử, mặc dầu chúng hoàn toàn có thể Giao hàng như thể vật tư di truyền dạng thô cho sự sản sinh gene mới trải qua quy trình nhân đôi ( gene duplication ) và phân ly gene. [ 104 ]
Phiên mã và dịch mã[sửa|sửa mã nguồn]
 Mã di truyền : DNA, qua trung gian RNA thông tin, mã hóa cho protein với những bộ ba mã hóa .
Mã di truyền : DNA, qua trung gian RNA thông tin, mã hóa cho protein với những bộ ba mã hóa .
Mỗi gene là một đoạn trình tự DNA chứa thông tin di truyền và có thể ảnh hưởng đến kiểu hình của sinh vật. Bên trong một gene, trình tự các base dọc theo một mạch DNA xác định nên trình tự của RNA thông tin, rồi từ đó xác lập nên trình tự của một hay nhiều protein. Mối liên hệ giữa trình tự nucleotide của các gene và trình tự các amino acid của protein được xác định bởi những quy tắc trong quá trình dịch mã, được biết đến với cái tên bộ mã di truyền. Mỗi mã di truyền chứa bộ ba ‘chữ cái’ gọi là triplet (bộ ba mã gốc) trên DNA hay codon (bộ ba mã sao) trên mRNA hay anticodon (bộ ba đối mã) trên tRNA tạo thành một trình tự gồm ba nucleotide (v.d. ACT, CAG, TTT trên mạch gốc DNA).
Trong quy trình phiên mã, những triplet của một gene được sao chép sang RNA thông tin thành những codon tương ứng bằng enzyme RNA polymerase. Bản sao RNA này sau đó được giải thuật bởi ribosome trải qua hoạt động giải trí đọc trình tự RNA bằng cách bổ trợ cặp base trong RNA thông tin với RNA luân chuyển, loại phân tử mang theo amino acid. Vì có bốn loại base khác nhau được tổ hợp thành những mã bộ ba, do vậy có toàn bộ 64 codon ( tổng hợp 43 ). Tất cả chúng được phân chia để mã hóa cho 20 loại amino acid cơ bản của sự sống, do đó một amino acid hoàn toàn có thể có nhiều hơn một codon mã hóa cho nó. Bên cạnh đó cũng có ba codon ‘ kết thúc ‘ hoặc ‘ không có ý nghĩa ‘ ( nonsense ) ghi lại điểm kết thúc của một vùng mã hóa ; chúng là những codon UAA, UAG và UGA ( tương ứng với những triplet TAA, TAG và TGA ) .
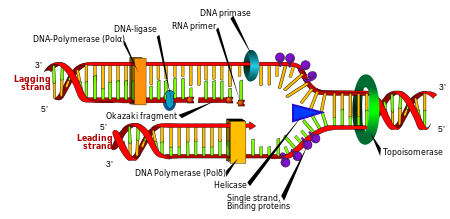 (leading strand) theo nguyên tắc bổ sung. Những DNA polymerase khác liên kết với mạch khuôn có chiều 5′ → 3′ ngược với chiều tháo xoắn tổng hợp nên (lagging strand) thành những đoạn ngắt quãng gọi là Nhân đôi DNA. Chuỗi xoắn kép được tháo xoắn theo chiều 3 ‘ → 5 ‘ bởi enzyme topoisomerase và cắt tách hai mạch đơn bởi enzyme helicase. Tiếp theo, một DNA polymerase lần lượt link liên tục những nucleotide tự do từ môi trường tự nhiên nội bào với những nucleotide trên mạch khuôn có chiều 3 ‘ → 5 ‘ tổng hợp nên mạch đứng vị trí số 1 theo nguyên tắc bổ trợ. Những DNA polymerase khác link với mạch khuôn có chiều 5 ‘ → 3 ‘ ngược với chiều tháo xoắn tổng hợp nên mạch theo sau thành những đoạn ngắt quãng gọi là đoạn Okazaki. Sau đó, những đoạn Okazaki này sẽ được nối lại với nhau bởi enzyme DNA ligase
(leading strand) theo nguyên tắc bổ sung. Những DNA polymerase khác liên kết với mạch khuôn có chiều 5′ → 3′ ngược với chiều tháo xoắn tổng hợp nên (lagging strand) thành những đoạn ngắt quãng gọi là Nhân đôi DNA. Chuỗi xoắn kép được tháo xoắn theo chiều 3 ‘ → 5 ‘ bởi enzyme topoisomerase và cắt tách hai mạch đơn bởi enzyme helicase. Tiếp theo, một DNA polymerase lần lượt link liên tục những nucleotide tự do từ môi trường tự nhiên nội bào với những nucleotide trên mạch khuôn có chiều 3 ‘ → 5 ‘ tổng hợp nên mạch đứng vị trí số 1 theo nguyên tắc bổ trợ. Những DNA polymerase khác link với mạch khuôn có chiều 5 ‘ → 3 ‘ ngược với chiều tháo xoắn tổng hợp nên mạch theo sau thành những đoạn ngắt quãng gọi là đoạn Okazaki. Sau đó, những đoạn Okazaki này sẽ được nối lại với nhau bởi enzyme DNA ligase
Nhân đôi DNA ( sao chép, tái bản )[sửa|sửa mã nguồn]
Phân bào là quy trình cơ bản của sinh vật để hoàn toàn có thể sinh trưởng, nhưng khi một tế bào phân loại, nó phải nhân đôi DNA trong bộ gene của nó sao cho hai tế bào con có cùng thông tin di truyền như của tế bào mẹ. Cấu trúc mạch kép DNA giúp hình thành một chính sách đơn thuần cho quy trình nhân đôi DNA. Ở đây, hai mạch đơn tháo xoắn tách rời nhau và mỗi mạch mới bổ trợ với mỗi mạch gốc được tổng hợp bằng một loại enzyme gọi là DNA polymerase. Enzyme này tạo ra những mạch mới bằng cách tìm những nucleotide tự do từ thiên nhiên và môi trường nội bào và kết nối đúng chuẩn với nucleotide trên mạch gốc bắt đầu theo nguyên tắc bổ trợ. Vì DNA polymerase chỉ tổng hợp mạch mới theo chiều 5 ′ → 3 ′, do vậy trên mạch khuôn có chiều 3 ‘ → 5 ‘ thì mạch bổ trợ được tổng hợp liên tục do cùng chiều với chiều tháo xoắn. [ 105 ] Còn trên mạch khuôn có chiều 5 ‘ → 3 ‘ thì mạch bổ trợ được tổng hợp ngắt quãng tạo nên những đoạn ngắn gọi là đoạn Okazaki do ngược chiều với chiều tháo xoắn, sau đó những đoạn này được nối lại với nhau nhờ enzyme nối DNA ligase. [ 106 ]
acid nucleic ngoại bào[sửa|sửa mã nguồn]
DNA ngoại bào trần (extracellular DNA – eDNA), hầu hết được giải phóng khi tế bào chết đi, xuất hiện khắp nơi trong môi trường. Mức độ tập trung của nó trong đất có thể lên tới 2 μg/lít, và trong môi trường nước tự nhiên lên tới 88 μg/lít.[107] Đã có một số chức năng khả thi của eDNA được đề xuất: nó có thể tham gia vào chuyển gene ngang;[108] cung cấp dinh dưỡng;[109] và có khả năng hoạt động như một chất đệm để khôi phục hoặc chuẩn độ ion hoặc tính kháng sinh.[110] DNA ngoại bào hoạt động như một thành phần chức năng của chất nền ngoại bào trong lớp màng vi sinh vật (phim sinh học – biofilm) của một số loài vi khuẩn. Nó có thể hoạt động như một nhân tố nhận diện để điều phối sự bám dính và phân tán của một số loại tế bào đặc hiệu trong phim sinh học;[111] hoặc đóng góp vào sự hình thành phim sinh học;[112] cũng như đóng góp vào đặc tính vật lý chắc chắn của phim sinh học và sức đề kháng trước những căng thẳng sinh học (biological stress).[113]
Tương tác với protein[sửa|sửa mã nguồn]
Mọi tính năng của DNA nhờ vào vào tương tác với protein. Những tương tác protein này hoàn toàn có thể không đặc hiệu hoặc đặc hiệu khi protein link với một trình tự DNA đơn cử. Các enzyme cũng link với DNA và trong số này, những enzyme polymerase sao chép trình tự base của DNA trong quy trình phiên mã và nhân đôi DNA có vai trò đặc biệt quan trọng quan trọng .
Protein link DNA[sửa|sửa mã nguồn]

Tương tác của DNA ( màu cam ) với protein histone ( màu lam ). Những amino acid cơ bản của protein link với những nhóm phosphat tính acid trên DNA .
Các protein cấu trúc liên kết với DNA là những ví dụ đã được nghiên cứu khá kĩ về tương tác không đặc hiệu DNA-protein. Bên trong nhiễm sắc thể, DNA được giữ trong phức hợp với protein cấu trúc. Những protein này tổ chức DNA thành một cấu trúc thắt đặc gọi là chất nhiễm sắc (chromatin). Trong sinh vật nhân thực, cấu trúc này bao gồm DNA liên kết với phức hợp các đơn vị protein cơ sở nhỏ gọi là histone, trong khi ở sinh vật nhân sơ lại có nhiều loại protein tham gia hơn.[114][115] Các histone tạo thành một phức hợp dạng đĩa gọi là nucleosome, với chuỗi xoắn kép DNA bao quanh bề mặt cấu trúc bằng hai vòng xoắn. Những tương tác không đặc hiệu được hình thành thông qua các phần dư cơ bản trong histone, tạo ra liên kết ion với bộ khung đường-phosphat có tính acid của DNA, và do vậy phần lớn tương tác là độc lập với trình tự các base.[116] Những phản ứng hóa học làm thay đổi các amino acid cơ bản này bao gồm phản ứng methyl hóa, phosphoryl hóa và acethyl hóa.[117] Những thay đổi hóa học này làm ảnh hưởng tới cường độ tương tác giữa DNA và histone, khiến cho các nhân tố phiên mã trở nên dễ dàng hoặc khó tiếp cận được với DNA và do vậy thay đổi tốc độ quá trình phiên mã.[118] Những protein liên kết DNA không đặc hiệu khác trong chất nhiễm sắc bao gồm các nhóm protein có tính linh động cao mà khi liên kết có thể uốn hoặc làm vặn DNA.[119] Các protein này có vai trò quan trọng trong việc sắp uốn nucleosome và xếp đặt chúng thành những cấu trúc lớn hơn tạo thành nhiễm sắc thể.[120]
Có một nhóm protein liên kết DNA đặc biệt là các protein chỉ liên kết đặc hiệu với một mạch đơn DNA. Ở người, protein A phục vụ quá trình nhân đôi DNA là protein được hiểu biết rõ ràng nhất trong nhóm này và tham gia vào những quá trình khi hai mạch xoắn kép đã tách rời nhau, bao gồm sao chép DNA, tái tổ hợp và sửa chữa DNA.[121] Những protein liên kết này giúp ổn định hóa mạch đơn DNA và bảo vệ nó khỏi hiện tượng hình thành cấu trúc vòng gấp kẹp tóc (stem-loop/hairpin loop) hoặc bị phân cắt bởi enzyme nuclease.
 (helix-turn-helix – HTH) gắn vào DNA đích.[122]Protein tác nhân ức chế phiên mã lambda motif cấu trúc xoắn-ngoặt-xoắn gắn vào DNA đích .
(helix-turn-helix – HTH) gắn vào DNA đích.[122]Protein tác nhân ức chế phiên mã lambda motif cấu trúc xoắn-ngoặt-xoắn gắn vào DNA đích .
Ngược lại, có những protein khác phải biến đổi cấu hình để liên kết với những trình tự DNA riêng biệt. Lĩnh vực nghiên cứu sâu rộng nhất về những protein này đó là nghiên cứu nhiều loại nhân tố phiên mã (transcription factor) khác nhau, đây chính là các protein điều hòa quá trình phiên mã. Mỗi nhân tố phiên mã liên kết với một tập hợp cụ thể các trình tự DNA và kích hoạt hoặc ức chế hoạt động phiên mã của gene tại những trình tự gần với vùng khởi động của chúng. Nhân tố phiên mã thực hiện vai trò này theo hai cách. Đầu tiên, chúng có thể gắn với RNA polymerase chịu trách nhiệm cho quá trình phiên mã, hoặc trực tiếp hoặc gián tiếp thông qua các protein trung gian; giúp định vị polymerase tại vùng gene khởi động và cho phép bắt đầu phiên mã.[123] Hoặc cách khác, nhân tố phiên mã có thể gắn với enzyme làm biến đổi các histone ở vùng khởi động. Điều này làm thay đổi khả năng tiếp cận của polymerase với mạch khuôn DNA.[124]
Do những DNA đích này Open trong toàn thể bộ gene sinh vật, thế cho nên những đổi khác trong hoạt động giải trí của một loại tác nhân phiên mã hoàn toàn có thể tác động ảnh hưởng tới hàng nghìn gene. [ 125 ] Hệ quả là, những protein này thường là tiềm năng của những quy trình truyền tín hiệu tải nạp ( signal transduction ) mà điều khiển và tinh chỉnh sự đáp ứng đối với những đổi khác của thiên nhiên và môi trường hoặc biệt hóa tế bào và tinh chỉnh và điều khiển sự tăng trưởng. Nét đặc trưng của những tương tác của những tác nhân phiên mã với DNA đến từ những protein tạo nhiều tiếp xúc với những cạnh của những base DNA, được cho phép chúng ” đọc ” được trình tự DNA. Phần lớn những tương tác với base diễn ra ở rãnh lớn, nơi hoàn toàn có thể tiếp xúc nhiều nhất với những base. [ 34 ]
Enzyme chỉnh sửa DNA[sửa|sửa mã nguồn]
Nuclease và ligase[sửa|sửa mã nguồn]
Nuclease là những enzyme có năng lực cắt mạch DNA bằng cách xúc tác cho phản ứng thủy phân những link phosphodieste. Loại nuclease thủy phân nucleotide từ những đầu mút của mạch DNA được gọi là exonuclease, trong khi endonuclease lại phân cắt từ những điểm trong mạch. Những nuclease được sử dụng liên tục nhất trong sinh học phân tử là những endonuclease số lượng giới hạn, do chúng cắt DNA tại những đoạn trình tự đặc hiệu. Ví dụ, enzyme EcoRV ở hình ảnh bên trái nhận ra trình tự gồm 6 base 5 ′ – GATATC-3 ′ và triển khai việc cắt theo một đường nằm ngang. Trong tự nhiên, những enzyme này bảo vệ vi trùng chống lại sự tiến công của thể thực khuẩn bằng cách tiêu hóa DNA thể thực khuẩn khi chúng xâm nhập vào tế bào vi trùng, lúc này những enzyme hoạt động giải trí như một phần trong mạng lưới hệ thống hạn chế cải biến ( restriction modification system ). [ 127 ] Trong công nghệ sinh học, những nuclease hoạt động giải trí với những trình tự đặc hiệu được sử dụng trong tách dòng phân tử ( molecular cloning ) và kỹ thuật nhận diện DNA ( DNA profiling ) .
 (replication fork):
(replication fork):
a: mạch khuôn, b: mạch dẫn đầu (leading strand), c: mạch theo sau (lagging strand), d: chạc tái bản, e: đoạn mồi RNA, f: các đoạn OkazakiMinh họa cấu trúc chạc tái bảna : mạch khuôn, b : mạch đứng vị trí số 1, c : mạch theo sau, d : chạc tái bản, e : đoạn mồi RNA, f : những đoạn OkazakiNhững enzyme có công dụng nối lại những đoạn DNA bị cắt hoặc bị đứt gãy được gọi là DNA ligase. [ 128 ] Ligase đặc biệt quan trọng quan trọng trong việc nối lại những mạch theo sau ngắt quãng của DNA, tức là những đoạn Okazaki tại chạc tái bản thành một bản sao hoàn hảo từ mạch khuôn DNA. Chúng cũng tham gia vào việc thay thế sửa chữa DNA và tái tổng hợp di truyền. [ 128 ]
Topoisomerase và helicase[sửa|sửa mã nguồn]
- Topoisomerase là những enzyme mang hoạt tính của cả nuclease lẫn ligase. Những protein này có khả năng thay đổi cấu trúc chuỗi xoắn kép DNA: chúng có thể thoái bỏ trạng thái siêu xoắn, hoặc ngược lại, chúng đóng xoắn. Một số enzyme trong nhóm này thực hiện hoạt động cắt chuỗi xoắn kép DNA và cho phép một phần phân tử quay được, do vậy làm giảm mức siêu xoắn của nó; sau cuối enzyme sẽ gắn khít hoàn chỉnh lại đoạn DNA bị gãy.[46] Những loại enzyme khác có thể cắt một chuỗi xoắn kép DNA và rồi kéo một mạch DNA thứ hai vào vị trí cắt này, trước khi thực hiện việc nối lại chuỗi xoắn kép.[129] Topoisomerase cần thiết cho nhiều quá trình liên quan đến DNA, như nhân đôi và phiên mã.[47]
- Helicase là những protein thuộc một trong những loại động cơ phân tử. Chúng sử dụng năng lượng hóa học trong nucleoside triphosphat, thường sử dụng nhất là adenosine triphosphat (ATP), để phá vỡ liên kết hydro giữa các base và tháo xoắn chuỗi kép DNA thành hai mạch đơn.[130] Những enzyme này có vai trò quan trọng thiết yếu đối với hầu hết quá trình enzyme cần thiết có tương tác với các base nitơ.
![]() Chrironumus pallidivitatus (chụp năm 2005).Ảnh chụp qua kính hiển vi điện tử của DNA : những đơn vị chức năng rRNA của ( chụp năm 2005 ) .
Chrironumus pallidivitatus (chụp năm 2005).Ảnh chụp qua kính hiển vi điện tử của DNA : những đơn vị chức năng rRNA của ( chụp năm 2005 ) .
Polymerase là những enzyme thực hiện tổng hợp mạch polynucleotide từ nucleoside triphosphat. Tính tuần tự của các sản phẩm của chúng được sinh ra dựa trên những mạch polynucleotide đã có—gọi là mạch khuôn. Những enzyme này hoạt động bằng lần lượt thêm vào một nucleotide tại nhóm 3′ hydroxyl ở điểm cuối của mạch polynucleotide đang phát triển. Kết quả là, mọi polymerase hoạt động luôn theo chiều từ đầu 5′ đến đầu 3′.[131] Tại trung tâm hoạt động của các enzyme này, phân tử nucleoside triphosphat đi đến ghép cặp với base của mạch khuôn: điều này cho phép polymerase tổng hợp một cách chính xác mạch bổ sung đối với mạch khuôn của nó. Các polymerase được phân loại theo các nhóm mạch khuôn mà chúng sử dụng.
Trong quy trình sao chép DNA, DNA polymerase phụ thuộc vào DNA tạo nên những bản sao của những mạch polynucleotide DNA. Để bảo toàn thông tin sinh học, điều cơ bản là trình tự của những base trong mỗi bản sao là trình tự bổ trợ đúng mực cho trình tự base trong mạch khuôn mẫu. Nhiều DNA polymerase có hoạt tính đọc và sửa sai ( proofreading ). Ở đây, polymerase nhận ra những lỗi thường Open trong phản ứng tổng hợp do sự thiếu đi những base ghép cặp giữa những nucleotide không khớp với nhau. Nếu polymerase phát hiện một sự không ăn khớp, hoạt tính exonuclease 3 ‘ – 5 ′ được kích hoạt và base không khớp nào được phát hiện sẽ bị cắt bỏ. [ 132 ] Trong hầu hết những sinh vật, DNA polymerase hoạt động giải trí trong một phức hệ lớn gọi là replisome có chứa nhiều tiểu đơn vị chức năng phụ, như protein kẹp DNA ( DNA clamp ) hay helicase. [ 133 ]DNA polymerase phụ thuộc vào RNA là những loại polymerase chuyên biệt triển khai sao chép trình tự của mạch RNA sang DNA. Chúng gồm có enzyme phiên mã ngược ( reverse transcriptase, RT ), ví dụ như một enzyme của virut retrovirus tham gia vào quy trình xâm nhập tế bào, và telomerase, cần cho quy trình sao chép telomere. [ 67 ] [ 134 ] Telomerase là một polymerase khác thường chính do nó chứa chính mạch khuôn RNA của nó như thể một phần trong cấu trúc của enzyme này. [ 68 ]
Sự phiên mã được thực hiện bởi RNA polymerase phụ thuộc DNA thông qua quá trình sao chép trình tự của mạch DNA sang RNA. Để bắt đầu giải mã một gene, RNA polymerase gắn với một trình tự của DNA gọi là vùng khởi động (promoter) và tách hai mạch DNA khỏi nhau. Sau đó nó sao chép trình tự gene vào một RNA thông tin cho đến khi nó đi đến vùng kết thúc (terminator) của DNA, nơi RNA polymerase dừng lại và tách khỏi DNA. Với DNA polymerase phụ thuộc DNA ở người, RNA polymerase II, enzyme thực hiện phiên mã hầu hết các gene trong bộ gene người, hoạt động như là một phần của một phức hệ protein lớn với nhiều tiểu đơn vị phụ và vùng điều hòa khác nhau.[135]
Tái tổng hợp di truyền[sửa|sửa mã nguồn]
 Tái tổng hợp gồm có tách rời và liên kết lại hai nhiễm sắc thể ( M và F ) để tạo thành hai nhiễm sắc thể được sắp xếp lại ( C1 và C2 ) .Chuỗi xoắn kép DNA thường không tương tác với những đoạn khác của DNA, và trong tế bào người những nhiễm sắc thể khác nhau thậm chí còn còn nằm ở những vùng tách biệt trong nhân tế bào gọi là ” vùng nhiễm sắc thể ” ( chromosome territory ). [ 137 ] Sự tách biệt về khoảng trống giữa những nhiễm sắc thể khác nhau là quan trọng so với năng lực hoạt động giải trí của DNA như là nơi lưu giữ không thay đổi thông tin di truyền, khi một vài lần nhiễm sắc thể tương tác trong sự trao đổi chéo nhiễm sắc thể xảy ra trong quy trình sinh sản hữu tính, khi ấy tái tổng hợp di truyền mới diễn ra. Trao đổi chéo nhiễm sắc thể là khi hai chuỗi DNA tháo xoắn và tách rời từng mạch đơn ra, trao đổi những đoạn DNA cho nhau rồi tái kết nối hai mạch đơn lại .Tái tổng hợp được cho phép nhiễm sắc thể trao đổi thông tin di truyền và tạo ra những tổng hợp gene mới, làm tăng hiệu suất cao của tính tinh lọc tự nhiên và hoàn toàn có thể quan trọng so với sự tiến hóa nhanh gọn cho những protein mới. [ 138 ] Tái tổng hợp di truyền cũng gồm có trong quy trình sửa chữa thay thế DNA, đặc biệt quan trọng trong sự phân phối của tế bào so với sự kiện chuỗi xoắn kép bị đứt gãy. [ 139 ]
Tái tổng hợp gồm có tách rời và liên kết lại hai nhiễm sắc thể ( M và F ) để tạo thành hai nhiễm sắc thể được sắp xếp lại ( C1 và C2 ) .Chuỗi xoắn kép DNA thường không tương tác với những đoạn khác của DNA, và trong tế bào người những nhiễm sắc thể khác nhau thậm chí còn còn nằm ở những vùng tách biệt trong nhân tế bào gọi là ” vùng nhiễm sắc thể ” ( chromosome territory ). [ 137 ] Sự tách biệt về khoảng trống giữa những nhiễm sắc thể khác nhau là quan trọng so với năng lực hoạt động giải trí của DNA như là nơi lưu giữ không thay đổi thông tin di truyền, khi một vài lần nhiễm sắc thể tương tác trong sự trao đổi chéo nhiễm sắc thể xảy ra trong quy trình sinh sản hữu tính, khi ấy tái tổng hợp di truyền mới diễn ra. Trao đổi chéo nhiễm sắc thể là khi hai chuỗi DNA tháo xoắn và tách rời từng mạch đơn ra, trao đổi những đoạn DNA cho nhau rồi tái kết nối hai mạch đơn lại .Tái tổng hợp được cho phép nhiễm sắc thể trao đổi thông tin di truyền và tạo ra những tổng hợp gene mới, làm tăng hiệu suất cao của tính tinh lọc tự nhiên và hoàn toàn có thể quan trọng so với sự tiến hóa nhanh gọn cho những protein mới. [ 138 ] Tái tổng hợp di truyền cũng gồm có trong quy trình sửa chữa thay thế DNA, đặc biệt quan trọng trong sự phân phối của tế bào so với sự kiện chuỗi xoắn kép bị đứt gãy. [ 139 ]
Dạng phổ biến nhất của trao đổi chéo nhiễm sắc thể là tái tổ hợp tương đồng, khi hai nhiễm sắc thể tham gia quá trình trên có trình tự DNA tương đồng. Tái tổ hợp không tương đồng có thể phá hủy tế bào, gây ra chuyển đoạn nhiễm sắc thể và biến dị di truyền. Phản ứng tái tổ hợp được xúc tác bởi các enzyme recombinase, như RAD51.[140] Bước đầu tiên trong quá trình tái tổ hợp là một chuỗi DNA bị đứt gãy do tác động bởi enzyme endonuclease hay những phá hủy đối với DNA.[141] Một loạt các bước tiếp theo có sự xúc tác một phần của recombinase, sau đó hai chuỗi xoắn kép nối lại tại ít nhất một điểm giao Holliday (Holliday junction), trong đó một đoạn của mạch đơn của chuỗi xoắn kép này được ghép nối với đoạn mạch đối ứng của chuỗi xoắn kép kia. Điểm giao Holliday là một cấu trúc tiếp xúc bốn nhánh mà có thể di chuyển dọc theo cặp nhiễm sắc thể, tráo đổi một mạch sang cho mạch khác. Phản ứng tái tổ hợp dừng lại khi điểm giao Holliday bị đứt và xảy ra quá trình hàn gắn lại chuỗi DNA được giải phóng.[142]
DNA chứa thông tin di truyền được cho phép toàn bộ dạng sống tân tiến hoạt động giải trí công dụng, sinh trưởng và sinh sản. Tuy nhiên, không rõ bao lâu trong hành trình dài lịch sử vẻ vang 4 tỷ năm của sự sống DNA đã khởi đầu tiếp đón tính năng này, vì có những yêu cầu cho rằng những dạng sống Open sớm nhất có năng lực đã sử dụng phân tử RNA thay vì DNA làm vật tư di truyền. [ 143 ] [ 144 ] RNA hoàn toàn có thể đã trở thành thành phần TT của quy trình trao đổi chất trong những tế bào sơ khai vì phân tử này hoàn toàn có thể vừa truyền đạt thông tin di truyền cũng như mang hoạt tính xúc tác phản ứng dưới dạng ribozyme. [ 145 ] Thế giới RNA cổ xưa này, một nơi acid nucleic được sử dụng cho cả quy trình xúc tác và di truyền, hoàn toàn có thể ảnh hưởng tác động đến sự tiến hóa của mạng lưới hệ thống mã di truyền hiện tại trên cơ sở bốn loại nucleobase. Điều này thực sự đã xảy ra, vì số lượng của những base khác nhau trong một khung hình sống như thể một sự thỏa hiệp giữa một số lượng nhỏ base tăng cường qua hoạt động giải trí nhân đôi đúng chuẩn và một số lượng lớn những base tăng cường qua hoạt động giải trí xúc tác hiệu suất cao của ribozyme. [ 146 ] Không may thay, trong thực tiễn lại không có vật chứng trực tiếp nào của mạng lưới hệ thống di truyền cổ xưa, như việc hồi sinh DNA từ phần đông những hóa thạch là điều không hề vì phân tử DNA chỉ sống sót trong thiên nhiên và môi trường ít hơn một triệu năm và từ từ phân hủy thành những mảnh ngắn tan vào dung dịch. [ 147 ] Những nhu yếu khảo sát so với dạng DNA cổ xưa đã được thực thi, trong đó báo cáo giải trình đáng quan tâm nhất là về sự cô lập của một loại vi trùng sống sót tăng trưởng độc lập từ một tinh thể muối có niên đại cách đây 250 triệu năm, [ 148 ] tuy nhiên những công bố này vẫn còn trong vòng tranh cãi. [ 149 ] [ 150 ]Những thành phần ” vữa gạch ” của DNA ( adenine, guanine và cả những phân tử hữu cơ tương quan ) hoàn toàn có thể đã hình thành từ thiên hà trong những khoảng chừng không liên thiên thể. [ 151 ] [ 152 ] [ 153 ] Những hợp chất hữu cơ cấu trúc nền tảng khác của DNA và RNA trong sự sống, gồm có uracil, cytosine và thymine, cũng đã được tổng hợp trong phòng thí nghiệm dưới những điều kiện kèm theo mô phỏng tương ứng tìm thấy trong khoảng trống ngoài thiên thể, bằng cách sử dụng những chất hóa học khởi đầu, ví dụ pyrimidine, tìm thấy trong những mảnh vẫn thạch. Pyrimidine, như những hydrocarbon đa vòng thơm ( polycyclic aromatic hydrocarbons – PAHs ), là hợp chất hóa học giàu carbon nhất tìm thấy trong thiên hà, hoàn toàn có thể được hình thành trong những ngôi sao 5 cánh khổng lồ đỏ hay trong những đám mây khí và bụi giữa những vì sao. [ 154 ]
Sử dụng trong công nghệ tiên tiến[sửa|sửa mã nguồn]
Kỹ thuật di truyền[sửa|sửa mã nguồn]
Nhiều giải pháp đã được tăng trưởng để sàng lọc DNA từ sinh vật sống, như chiết lỏng-lỏng phenol-clorofom, và vận dụng trong phòng thí nghiệm, như giải pháp phân hủy số lượng giới hạn ( restriction digest ) và phản ứng chuỗi polymerase. Sinh học văn minh và ngành hóa sinh sử dụng liên tục những kỹ thuật này trong công nghệ tiên tiến tái tổng hợp DNA. Tái tổng hợp DNA là một trình tự DNA nhân tạo được lắp ghép từ những trình tự DNA khác. Chúng hoàn toàn có thể được biến nạp vào tế bào sinh vật dưới dạng plasmid hoặc trong những dạng thích hợp khác, bằng cách sử dụng vector virut. [ 155 ] Những sinh vật biến hóa di truyền hoàn toàn có thể được ứng dụng để sinh ra những loại sản phẩm như protein tái tổng hợp, sử dụng trong điều tra và nghiên cứu y học, [ 156 ] hoặc được nuôi trồng trong nông nghiệp. [ 157 ] [ 158 ]
Kỹ thuật nhận diện DNA[sửa|sửa mã nguồn]
Các nhà khoa học pháp y sử dụng DNA trong máu, tinh dịch, da, nước bọt hay tóc tìm thấy tại hiện trường để nhận ra DNA khớp với của một cá nhân, như của thủ phạm chẳng hạn. Quá trình này được gọi là kỹ thuật nhận diện DNA (DNA profiling), hay còn gọi là kỹ thuật in dấu DNA (DNA fingerprintin)). Trong kỹ thuật nhận diện DNA, độ dài của nhiều đoạn DNA lặp lại, như các đoạn trình tự vi vệ tinh (microsatellite) và vệ tinh nhỏ (minisatellite), được so sánh giữa các cá nhân có liên quan. Phương pháp này thường là một kỹ thuật cực kỳ tin cậy cho phép xác định những trình tự DNA ăn khớp với nhau.[159] Tuy vậy, việc nhận dạng có thể trở nên phức tạp nếu tại hiện trường gây án có nhiều DNA của nhiều người.[160] Kỹ thuật nhận diện DNA phát triển vào năm 1984 bởi nhà di truyền học người Anh Sir Alec Jeffreys,[161] và lần đầu tiên được sử dụng trong ngành pháp y để cáo buộc Colin Pitchfork trong vụ án Enderby năm 1988.[162]
Sự tăng trưởng của khoa học pháp y, và năng lực lúc bấy giờ hoàn toàn có thể nhận ra thông tin di truyền từ những mẫu máu, da, nước bọt hay tóc đã dẫn đến nhiều vụ án phải lật lại hồ sơ mặc dầu tòa đã tuyên án. Chứng cứ mà lúc bấy giờ hoàn toàn có thể được bật mý ra trong khi ở thời gian thẩm vấn là bất khả thi về mặt khoa học. Kết hợp với luật đạo vô hiệu trường hợp bất trùng khả tố ( double jeopardy-một người không bị xử hai lần về một tội ) ở 1 số ít nơi, đã được cho phép khởi tố lại 1 số ít vụ án khi bản án trước đó đã không nêu được chứng cứ thuyết phục để phán quyết. Những người mang tội danh nặng được phép nhu yếu lấy mẫu DNA nhằm mục đích mục tiêu so sánh. Trường hợp biện hộ rõ ràng nhất đó là mẫu DNA nhận được từ pháp y bị cho là đã bị ảnh hưởng tác động từ những người ở xung quanh vụ án. Điều này làm cho những thủ tục tìm hiểu trở nên ngặt nghèo hơn trong những trường hợp phạm tội mới. Nhận diện DNA cũng được vận dụng thành công xuất sắc cho nhận dạng những nạn nhân trong những vụ tai nạn đáng tiếc có thương vong lớn, [ 163 ] từ những phần khung hình, và phân biệt từng nạn nhân trong những mồ chôn tập thể trong cuộc chiến tranh, trải qua so sánh với DNA của người nhà nạn nhân .Kỹ thuật nhận diện DNA cũng được sử dụng để xác nhận mối liên hệ sinh học với cha mẹ hoặc ông bà của một đứa trẻ với Xác Suất đúng chuẩn lên tới 99,99 %. Những giải pháp kiểm trình tự DNA thông thường được thực thi sau sinh, nhưng những chiêu thức mới hoàn toàn có thể kiểm tra quan hệ huyết thống ngay cả khi người mẹ đang mang thai. [ 164 ]
DNA enzyme hay xúc tác DNA[sửa|sửa mã nguồn]
Xem thêm thông tin : DeoxyribozymeDeoxyribozyme, cũng gọi là DNAzyme hay xúc tác DNA phát hiện lần tiên phong vào năm 1994. [ 165 ] Phần lớn chúng là những trình tự mạch đơn DNA được cô lập khỏi một vũng lớn gồm nhiều trình tự DNA ngẫu nhiên trải qua một hướng tiếp cận tổng hợp gọi là kỹ thuật lựa chọn in vitro hay giải pháp SELEX. Những DNAzyme tham gia xúc tác những phản ứng hóa học gồm có phân cắt RNA-DNA, liên kết RNA-DNA, sự phosphoryl hóa – phản phosphoryl hóa những amino acid, hình thành link carbon-carbon, v.v… DNAzyme hoàn toàn có thể tăng cường vận tốc phản ứng hóa học gấp 100.000.000.000 lần so với phản ứng không có sự tham gia xúc tác của nó. [ 166 ] Các DNAzyme được nghiên cứu và điều tra nhiều nhất là những loại phân cắt RNA dùng để phát hiện những ion sắt kẽm kim loại khác nhau và phong cách thiết kế những tác nhân trị liệu. Một vài DNAzyme đặc hiệu ion sắt kẽm kim loại gồm có GR-5 DNAzyme ( đặc hiệu với chì ), [ 165 ] CA1-3 DNAzymes ( với đồng ), [ 167 ] 39E DNAzyme ( với ion uranyl ) và NaA43 DNAzyme ( với natri ). [ 168 ] NaA43 DNAzyme, nhạy với natri gấp 10.000 lần so với những ion sắt kẽm kim loại khác, được dùng để theo dõi natri theo thời hạn thực trong tế bào sống .
Tin sinh học[sửa|sửa mã nguồn]
 Bản đồ nhiễm sắc thể X ở người (từ trang web của ( từ website của NCBI ) .Tin sinh học gồm có những kỹ thuật tàng trữ, khám phá tài liệu, tìm kiếm và thao tác với tài liệu sinh học, gồm có tài liệu về trình tự acid nucleic DNA. Các kỹ thuật này mang đến những ứng dụng thoáng đãng trong khoa học máy tính, đặc biệt quan trọng là thuật toán tìm kiếm chuỗi, học máy và kim chỉ nan cơ sở tài liệu. [ 169 ] Thuật toán tìm kiếm chuỗi hay so khớp, trong đó tìm kiếm sự Open của một trình tự những vần âm trong một trình tự những vần âm lớn hơn, được tăng trưởng để tìm ra những trình tự nucleotide đơn cử. [ 170 ] Trình tự DNA hoàn toàn có thể sắp gióng với những trình tự DNA khác để nhận ra những trình tự tương đương và xác lập vị trí đột biến khiến chúng độc lạ. Những kỹ thuật này, đặc biệt quan trọng là kỹ thuật ” sắp gióng đa trình tự ” ( multiple sequence alignment ), được sử dụng để điều tra và nghiên cứu những mối quan hệ phát sinh chủng loài học và tính năng của protein. [ 171 ] Tập hợp tài liệu của hàng loạt trình tự DNA, như được lập ra bởi Dự án map gene người, là khó để sử dụng mà không có những chú giải được cho phép nhận ra vị trí của những gene hay những yếu tố điều hòa ở mỗi nhiễm sắc thể. Vùng trình tự DNA với những phần đặc trưng gắn với gene mã hóa cho protein hoặc RNA hoàn toàn có thể tìm ra bằng thuật toán tìm kiếm gene ( gene finding algorithm ), được cho phép những nhà nghiên cứu Dự kiến sự xuất hiện của những sinh phẩm đặc biệt quan trọng mã hóa bởi gen và công dụng của chúng trong sinh vật trước khi chúng được phát hiện bằng thực nghiệm. [ 172 ] Toàn bộ hệ gene cũng hoàn toàn có thể được đối sánh tương quan, để làm sáng tỏ lịch sử vẻ vang tiến hóa của từng sinh vật đơn cử và được cho phép kiểm tra những sự kiện tiến hóa phức tạp .
Bản đồ nhiễm sắc thể X ở người (từ trang web của ( từ website của NCBI ) .Tin sinh học gồm có những kỹ thuật tàng trữ, khám phá tài liệu, tìm kiếm và thao tác với tài liệu sinh học, gồm có tài liệu về trình tự acid nucleic DNA. Các kỹ thuật này mang đến những ứng dụng thoáng đãng trong khoa học máy tính, đặc biệt quan trọng là thuật toán tìm kiếm chuỗi, học máy và kim chỉ nan cơ sở tài liệu. [ 169 ] Thuật toán tìm kiếm chuỗi hay so khớp, trong đó tìm kiếm sự Open của một trình tự những vần âm trong một trình tự những vần âm lớn hơn, được tăng trưởng để tìm ra những trình tự nucleotide đơn cử. [ 170 ] Trình tự DNA hoàn toàn có thể sắp gióng với những trình tự DNA khác để nhận ra những trình tự tương đương và xác lập vị trí đột biến khiến chúng độc lạ. Những kỹ thuật này, đặc biệt quan trọng là kỹ thuật ” sắp gióng đa trình tự ” ( multiple sequence alignment ), được sử dụng để điều tra và nghiên cứu những mối quan hệ phát sinh chủng loài học và tính năng của protein. [ 171 ] Tập hợp tài liệu của hàng loạt trình tự DNA, như được lập ra bởi Dự án map gene người, là khó để sử dụng mà không có những chú giải được cho phép nhận ra vị trí của những gene hay những yếu tố điều hòa ở mỗi nhiễm sắc thể. Vùng trình tự DNA với những phần đặc trưng gắn với gene mã hóa cho protein hoặc RNA hoàn toàn có thể tìm ra bằng thuật toán tìm kiếm gene ( gene finding algorithm ), được cho phép những nhà nghiên cứu Dự kiến sự xuất hiện của những sinh phẩm đặc biệt quan trọng mã hóa bởi gen và công dụng của chúng trong sinh vật trước khi chúng được phát hiện bằng thực nghiệm. [ 172 ] Toàn bộ hệ gene cũng hoàn toàn có thể được đối sánh tương quan, để làm sáng tỏ lịch sử vẻ vang tiến hóa của từng sinh vật đơn cử và được cho phép kiểm tra những sự kiện tiến hóa phức tạp .
Công nghệ nano DNA[sửa|sửa mã nguồn]
Công nghệ nano DNA sử dụng những tính chất tương tác của phân tử DNA và những acid nucleic khác để tạo ra những phức hợp DNA phân nhánh tự lắp ráp có tính năng hữu ích.[173] Do vậy DNA được sử dụng như là vật liệu cấu trúc hơn là vật liệu mang thông tin sinh học. Các nhà khoa học đã tạo ra những dàn lưới hai chiều tuần hoàn (bằng phương pháp lát gạch và origami DNA) và cấu trúc ba chiều đa diện đều.[174] Thiết bị cơ nano và thuật toán tự lắp ráp cũng được chứng minh là khả dĩ,[175] và những cấu trúc DNA này dùng làm khuôn mẫu để sắp xếp các phân tử khác như keo vàng (colloidal gold) và protein streptavidin trong vi khuẩn Streptomyces avidinii.[176]
Lịch sử và nhân chủng học[sửa|sửa mã nguồn]
Bởi vì theo thời hạn DNA tích góp những đột biến, do vậy chúng được di truyền lại, nên DNA chứa thông tin lịch sử vẻ vang, và bằng cách so sánh những trình tự DNA, những nhà di truyền học hoàn toàn có thể suy luận ra lịch sử dân tộc tiến hóa của mỗi loài sinh vật, hay phát sinh chủng loài của chúng. [ 177 ] Lĩnh vực phát sinh chủng loài học là một công cụ mạnh của sinh học tiến hóa. Nếu so sánh những trình tự DNA của một loài với nhau, những nhà di truyền quần thể hoàn toàn có thể biết được lịch sử dân tộc tăng trưởng của một quần thể đang nghiên cứu và điều tra. Kết quả nghiên cứu và điều tra của ngành này được vận dụng sang cho di truyền sinh thái xanh và nhân chủng học. Ví dụ, những nhà khoa học đã sử dụng vật chứng DNA để điều tra và nghiên cứu sự kiện Mười bộ tộc biến mất ( Ten Lost Tribes ) của Israel. [ 178 ] [ 179 ]
Lưu trữ thông tin[sửa|sửa mã nguồn]
Trong một bài báo trên tạp chí Nature tháng 1 năm 2013, các nhà khoa học từ Học viện Tin sinh học Châu Âu và công ty Agilent Technologies đã đề xuất một cơ chế sử dụng khả năng mã hóa thông tin của DNA để phục vụ cho việc lưu trữ kỹ thuật số. Nhóm nghiên cứu mã hóa 739 kilobyte dữ liệu vào mã DNA, rồi tổng hợp nên DNA thực thụ, tiếp đó thực hiện giải trình tự DNA và giải mã thông tin ngược trở lại dạng ban đầu, mà họ thông báo là kết quả thu được với độ chính xác 100%. Thông tin được mã hóa chứa các tập tin định dạng văn bản và âm thanh. Một thí nghiệm khác thực hiện trước đó bởi nhóm nghiên cứu ở Đại học Harvard tháng 8 năm 2012, khi nhóm này mã hóa một quyển sách chứa 54.000 từ vào DNA.[180][181]
Trong tế bào sinh vật sống, thông tin lưu trữ ở DNA có thể được kích hoạt bởi các enzyme. Ví dụ như các kênh ion có protein cảm thụ ánh sáng phối hợp với enzyme xử lý DNA là phù hợp cho nhiệm vụ trên trong ống nghiệm (in vitro).[182][183] Những phân tử exonuclease huỳnh quang có thể truyền tín hiệu ra bên ngoài tuân theo các trình tự nucleotide mà chúng đọc được.[184]
Lịch sử điều tra và nghiên cứu DNA[sửa|sửa mã nguồn]
 Phác thảo bằng bút chì về chuỗi xoắn kép DNA do Francis Crick vẽ năm 1953 .DNA lần tiên phong được cô lập bởi thầy thuốc người Thụy Sĩ Friedrich Miescher, người mà vào năm 1869, đã tò mò ra một chất vi mô trong mủ của băng gạc được tháo bỏ sau phẫu thuật. Vì nó nằm trong nhân của tế bào, ông đã gọi nó là ” nuclein “. [ 185 ] [ 186 ] Năm 1878, Albrecht Kossel đã cô lập được thành phần không phải là protein của ” nuclein “, acid nucleic, và sau đó ông cô lập được năm nucleobase cơ bản của nó. [ 187 ] [ 188 ] Năm 1919, Phoebus Levene phân biệt được những đơn vị chức năng của nucleotide là base, đường và phosphat. [ 189 ] Levene đề xuất kiến nghị rằng DNA chứa một chuỗi những đơn vị chức năng nucleotide được link với nhau bằng những nhóm phosphat. Levene đã nghĩ rằng mạch này là ngắn và những base lặp lại theo một thứ tự cố định và thắt chặt. Năm 1937, William Astbury chụp được ảnh thành phần nhiễu xạ tia X tiên phong cho thấy DNA có một cấu trúc đều đặn. [ 190 ]
Phác thảo bằng bút chì về chuỗi xoắn kép DNA do Francis Crick vẽ năm 1953 .DNA lần tiên phong được cô lập bởi thầy thuốc người Thụy Sĩ Friedrich Miescher, người mà vào năm 1869, đã tò mò ra một chất vi mô trong mủ của băng gạc được tháo bỏ sau phẫu thuật. Vì nó nằm trong nhân của tế bào, ông đã gọi nó là ” nuclein “. [ 185 ] [ 186 ] Năm 1878, Albrecht Kossel đã cô lập được thành phần không phải là protein của ” nuclein “, acid nucleic, và sau đó ông cô lập được năm nucleobase cơ bản của nó. [ 187 ] [ 188 ] Năm 1919, Phoebus Levene phân biệt được những đơn vị chức năng của nucleotide là base, đường và phosphat. [ 189 ] Levene đề xuất kiến nghị rằng DNA chứa một chuỗi những đơn vị chức năng nucleotide được link với nhau bằng những nhóm phosphat. Levene đã nghĩ rằng mạch này là ngắn và những base lặp lại theo một thứ tự cố định và thắt chặt. Năm 1937, William Astbury chụp được ảnh thành phần nhiễu xạ tia X tiên phong cho thấy DNA có một cấu trúc đều đặn. [ 190 ]
Năm 1927, Nikolai Koltsov đề xuất rằng các tính trạng di truyền có thể được thừa hưởng thông qua một “phân tử di truyền khổng lồ” cấu thành từ “hai mạch đối xứng mà có thể sao chép theo cách bán bảo tồn sử dụng từng mạch như là một khuôn mẫu”.[191][192] Năm 1928, Frederick Griffith trong thí nghiệm của ông đã khám phá ra các tính trạng dạng “trơn” của phế cầu khuẩn (Pneumococcus) có thể truyền sang dạng “thô” của cùng một loài vi khuẩn bằng cách trộn các vi khuẩn dạng “trơn” đã bị giết với các vi khuẩn dạng “thô” còn sống bằng thí nghiệm nổi tiếng gọi là thí nghiệm Griffith.[193][194] Hệ thống thí nghiệm này cung cấp gợi ý rõ ràng đầu tiên về DNA mang thông tin di truyền—theo thí nghiệm Avery–MacLeod–McCarty—khi Oswald Avery, cùng với các đồng nghiệp Colin MacLeod và Maclyn McCarty, nhận ra DNA tuân theo nguyên lý biến nạp trong thí nghiệm Griffith vào năm 1943.[195] Vai trò của DNA trong di truyền được xác nhận vào năm 1952, khi Alfred Hershey và Martha Chase trong thí nghiệm Hershey–Chase chỉ ra rằng DNA là vật liệu di truyền của thực khuẩn thể T2.[196]
Năm 1953, James Watson và Francis Crick lần đầu tiên đề xuất mô hình mà được chấp nhận ngày nay với cấu trúc DNA chuỗi xoắn kép đăng trên tạp chí Nature.[12] Mô hình phân tử chuỗi xoắn kép DNA của họ khi ấy dựa trên ảnh chụp nhiễu xạ tia X (còn gọi là “Ảnh chụp 51”)[197] do Rosalind Franklin và Raymond Gosling thực hiện vào tháng 5 năm 1952, và dựa trên thông tin rằng các base DNA ghép cặp với nhau.
Chứng cứ thực nghiệm ủng hộ mô hình Watson và Crick được công bố trong một loạt 5 bài báo đăng trên cùng một số của tờ Nature.[198] Trong các bài báo này, bài viết của Franklin và Gosling là công trình đầu tiên của chính họ công bố dữ liệu về nhiễu xạ tia X và phương pháp phân tích gốc giúp ủng hộ một phần mô hình của Watson và Crick;[50][199] trong số báo này cũng bao gồm bài viết về cấu trúc DNA của Maurice Wilkins với hai đồng nghiệp của ông, khi họ thực hiện phân tích ảnh chụp tia X của dạng B-DNA trong cơ thể sống (in vivo) mà cũng ủng hộ cho sự có mặt trong cơ thể sống của cấu trúc chuỗi xoắn kép DNA như đề xuất của Crick và Watson về mô hình phân tử DNA của họ trong bài báo dài 2 trang đăng ở số trước của tạp chí Nature.[51] Năm 1962, khi ấy Franklin đã qua đời, Watson, Crick, và Wilkins cùng nhận Giải Nobel Sinh lý và Y học.[200] Do điều lệ của Quỹ Nobel chỉ trao giải cho những nhà khoa học còn sống. Vẫn có những tranh luận về sau liên quan đến những ai xứng đáng được công nhận liên quan đến khám phá này.[201]
Trong một buổi chuyện trò có tầm tác động ảnh hưởng vào năm 1957, Crick đưa ra luận thuyết TT của sinh học phân tử, báo hiệu trước về mối quan hệ giữa những phân tử DNA, RNA, và protein, và khớp nối với ” giả thuyết về dòng thông tin “. [ 202 ] Chứng cứ thực nghiệm ở đầu cuối xác nhận chính sách sao chép mà hàm ý cấu trúc chuỗi xoắn kép được công bố vào năm 1958 trải qua thí nghiệm Meselson – Stahl. [ 203 ] Những khu công trình về sau của Crick và những đồng nghiệp cũng như của nhiều nhà khoa học khác chứng tỏ mã di truyền có cơ sở là tổng hợp của bộ ba base không chồng lợp nhau, hay còn gọi là những codon, được cho phép Har Gobind Khorana, Robert W. Holley và Marshall Warren Nirenberg làm sáng tỏ mã di truyền. [ 204 ] Những phát hiện này đã khai sinh ra ngành sinh học phân tử .
Liên kết ngoài[sửa|sửa mã nguồn]
Source: https://blogchiase247.net
Category: Hỏi Đáp





